
Archaic points of potential interest
See page 194 in Schultes & Hofmann 1980 concerning the claims of Adavasio & Fry 1976 related to archeological finds of peyote in rock shelters and caves in the Cuatro Cienegas Basin in Coahuila, Mexico.
This is a fascinating article that is often mentioned in conjunction with their proposal that, over the millennia, sacramental plants were progressively replaced by safer ones. While it is a tantalizing notion to entertain, it needs to be understood that Adavasio & Fry did not include adequate evidence to support either of their assertions a) that the use of Ungnadia preceeded that of Sophora and b) both were eventually replaced with Lophophora. Three points need to be made:
First, any evidence presented for the sacramental use of the Mexican Buckeye (Ungnadia speciosa Endl.) is circumstantial at best, being primarily based on its form of packaging being similar to that of the Texas mountain-laurel (Sophora secundiflora (Ort.) Lag. ex SD), their occasional comingling, and was bolstered with only one second-hand reference to it being a euphoric plant. There does not appear to be any accounts of or archaeological evidence for it actually being used for human ingestion ceremonially. This is not a small point to overlook.

Ungnadia speciosa leaves

Ungnadia speciosa seeds
Second, Sophora itself is not known to have ever actually been used as a sacramental hallucinogen per se. It was employed in a highly dangerous form of a vision quest to gain animal spirits as guardians, protectors or familiars. Granted, it was incorporated into the peyote drink as prepared by several tribes and it is reasonable to assume that to be an ancient practice. It has been closely associated physically with peyote rites, strung as beads, as well as reported to have been used medicinally, ritually and as a “narcotic” (a word that has been so misapplied, misconstrued and misused anthropologically as to have little, if any, reliably discernible meaning).

Sophora secundiflora

Sophora secundiflora
I do not dismiss Sophora‘s sacred stature, nor would I want to dispute its long association with the peyote ritual but it is important to keep in mind that its use (or effectiveness) as a hallucinogen has never been adequately demonstrated. It certainly is not supported by any of the published human bioassays involving either the plant or its pure alkaloids.
Descriptions of its effects were given years ago, in an upper division University of Texas at Austin anthropology class taught by Dr. William W. Newcomb (ANT 322M: “Indians of Texas”). Newcomb presented the stage of interaction with animal spirits to occur while the subject was in a deep coma-like stupor. Their successful entry into this state was said to demonstrated to onlookers by their unresponsiveness to a toothy gar-fish jaw being raked across their flesh. After intense headache, profuse vomiting and violent convulsions, a “coma” ensued that was said to last for several days. [That commentary is not included in his book Indians of Texas but rather was taken directly from my class notes.]

Sophora secundiflora
Dr. Newcomb clearly presented it as being used ritually but not as a ritual sacrament or hallucinogen in the sense of peyote.
[See Schultes & Hofmann 1980 & 1992 for a broader discussion and also Hatfield et al. 1977 for chemical study and a discussion of the reports of ethnological use. LaBarre 1975 also includes a fascinating discussion.]
Ott 1993 mentions that despite claims by a number of people that the use of the ‘Red bean’ was the predecessor to and replaced by that of peyote, this was thoroughly challenged in a monograph by Merrill 1977.
I have had some very curious interactions with this plant; none of which involved its ingestion. It is definitely able to interact with people, and to a surprising degree to direct and influence their behavior to gather its seeds. I also believe it to be a sentient being based on my experiences with it as a living plant. It is not surprising that it was highly venerated by any people who knew it. It is also not surprising that they would gather its seeds in large numbers and store them in a special context. I’ve noticed that this still occurs although I am guessing modern containers of beans are maintained entirely for their decorative value.
Finally, my main objection to the conclusions of Adavasio & Fry concerning peyote is that they only selectively mention one solitary occurrence of the plant found by an earlier worker [Note 30].
It is very peculiar to attempt the support of such important conclusions with a single data point. Perhaps they also encountered additional examples but did not mention the details in this article. It is a puzzling omission considering their core assertion was one of safer plants being sequentially adopted.
Campbell 1958 had flatly asserted that it was not possible to demonstrate the priority of mescal beans over peyote based on the available evidence so something more is required.
It might also be added that there is a lack of evidence suggesting a continuity of culture in which a sequential series of sacramental replacements *could* occur but this seems trivial by comparison to items 1, 2 or 3.
No futher evidence was presented by Adavasio & Fry.
Disturbingly, the only find of peyote mentioned by Advasio & Fry in the same context as Sophora and Ungnadia was that of ONE string of dried peyote buttons dating from several millennia after peyote is known archeologically to have been employed by humans.
[See comments in Boyd & Dering 1996 concerning Ungnadia.]
The omission by Adovasio & Fry of any reference to additional finds is puzzling but it is far from being the only instance of cherry-picking that one can encounter in this area.
Schultes & Hofmann 1992 commented that peyote has been thought to have been in use for at least 4000 years. Dry cave and rock shelter finds in Texas are also said to have yielded dried samples of peyote that were three thousand years old and one find was purportedly dated to 7000 years. Those refer to specimens in the Witte Museum that have been claimed to be of that immense age (more on this in a moment.)
Oddly, until recently the entirety of the radiocarbon dating surrounding peyote has almost been characterized by missing information.
Let’s take a look at what is known concerning ancient peyote dating.
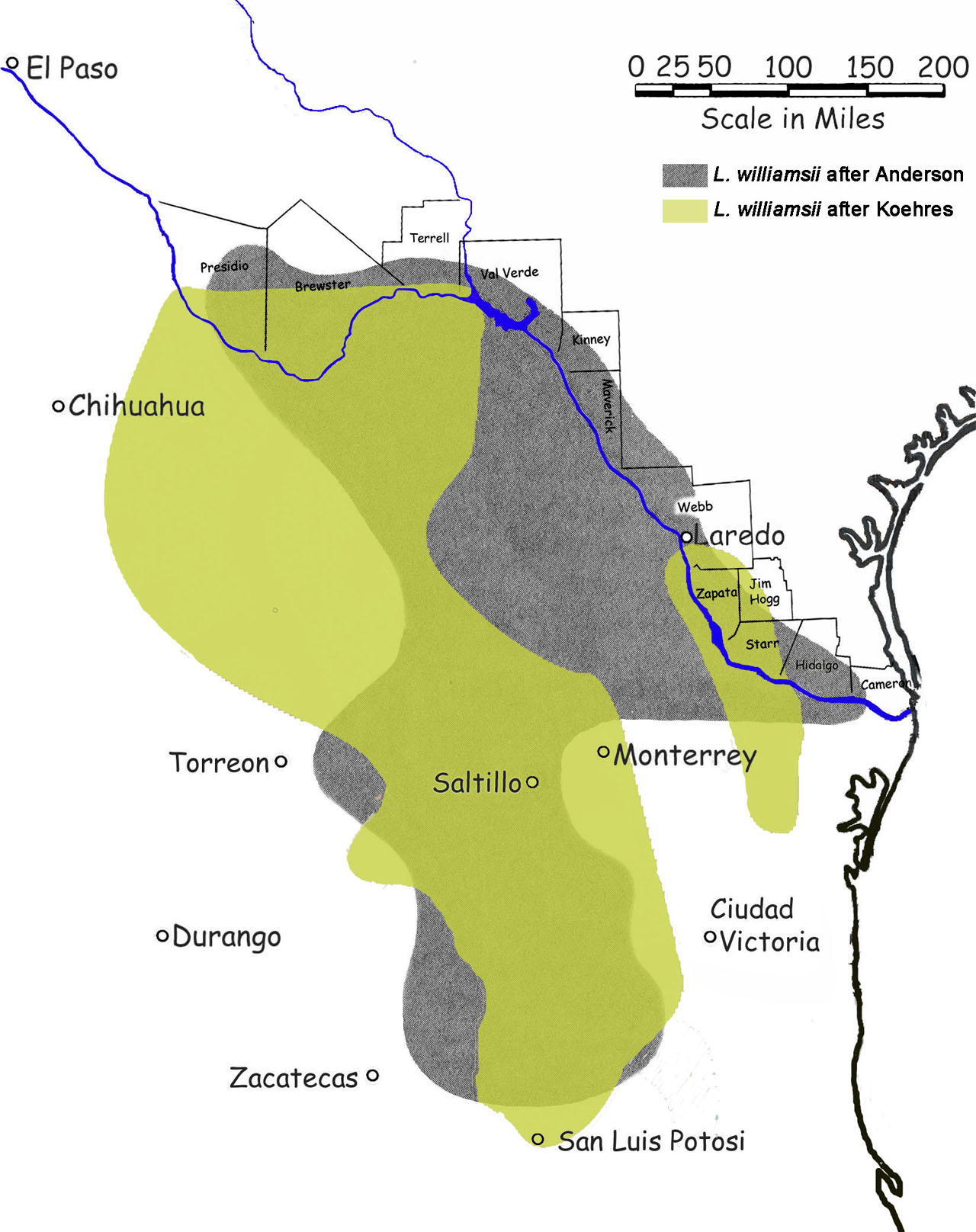
There are five reports where peyote was mentioned as being recovered from archaeological sites but material for only two of these can be located within the known collections. (See Terry et al. 2006 for details and references.)
One of these was from Cuatro Ciénegas, Coahuila, Mexico. This is a burial site from the transition between the Late Archaic and the Late Prehistoric Periods.
The account of Bruhn et al. 1978 gave its age based not on the actual dating of the peyote but on the dating of material thought to be associated with that single set of 8 strung and remarkably well preserved peyote buttons. Three mats had been dated with a range of 810-1070 AD (uncorrected values).
Due to the burial being a secondary interment, Martin Terry dated one of the actual Cuatro Ciénegas’ peyote specimens establishing its age as 835 ± 35 14C years BP.
Shumla Cave (No. 5, 41VV113) in southwest Texas on the other hand was an inhabited residential site (with intrusion of several burials.) The peyote was believed to have been deposited in the Eagle Nest subperiod of the Middle Archaic Period.
The exact provenience of the materials removed from the caves was not recorded but it is clear that they did mention recovering “a single mummified example” from Shumla Cave No. 5. This later became mistakenly presented as “petrified”. There was clearly more than one such artifact recovered from those excavations as three remained at the Witte for the removal of samples following the consumption of several others for earlier radiocarbon analysis. Terry et al. 2006 also reported their dating of the Shumla Caves’ peyote to 5195 ± 20 14C years BP.
Summary of earlier dating accounts:
Peter Furst was the first to report a date of 7000 BP for the Shumla cave material but due to several reasons the actual test data is either lost or irretrievable. It is therefore not possible to know anything further about the work or the claim. That claim oddly only appeared as a passing mention within a book review written by Furst. His date was repeated by Schultes & Hofmann and now appears stated as a fact throughout the literature.
Bruhn et al. 2002 presented a letter to the editor of Lancet in which they asserted establishing peyote use for 5700 years after dating the Shumla material (Furst’s 7000 BP). They repeated the claims in DeSmet & Bruhn 2003 citing Bruhn et al. 2002 as their primary reference.
El-Seedi et al. 2005 reported dating the Shumla Caves’ material to 3780-3660 BC.
Jan Bruhn (in a personal communication with Martin Terry) reported this was a weighted mean of 4952 ± 44 14C years.
Further investigation by Terry suggests that the discrepancy was most likely due to a failure of Bruhn to remove residual humic acid prior to radiocarbon dating. (See Terry et al. 2006)
Before leaving the subject of Sophora and Ungnadia completely, one point which must be considered is that world wide people have often venerated plants known to be deadly or exceedingly dangerous, with no intentions of using them for ingestion unless perhaps for the purpose of inducing death, or sometimes when ascertaining the guilt or innocence of a person by ordeal poisoning.
Many other ritual purposes besides hallucinogen use exist for plants. As the authors point out, the quantities found far exceed what would be needed for ingestion purposes. [Issue might be taken with their regarding a “crazed” state as being one conducive to, much less equated with, any ritual or sacred act. If anything it reflects a common culture-centric dismissal of what is viewed as a “more primitive” mindset.]
The most common usage of the red bean is that of ornamentation in the form of beads. The Mexican Buckeye would also make fine beads although none were mentioned as such.
Apparently the finds of both seeds consisted primarily of small caches of them, sometimes mixed together, which were sealed in plaited or twilled baskets which had to be torn to access the contents. [Both seeds also make a very nice sound, with good high frequency components, when shook or stirred. In quantity; the sound is absolutely mesmerizing. Such containers might have been musical instruments?]
In some sites, Sophora seeds and pods were found scattered throughout the cultural deposits.

Sophora secundiflora seedpods
Those twilled containers could also represent power objects used just as they were. We know between almost nothing and nothing about these people’s religious and spiritual beliefs. Some people who later used the beans believed they needed to be roasted until yellow, or forcefully struck and crushed, to ‘kill’ them before use. Since the seeds recovered in those finds were apparently being stored intact perhaps they were intended to assure protection after death, or at least to be available after death.
It may be noteworthy however that in the Murrah Cave many of the individual Sophora seeds were described by Holden 1937 as being “parched”; perhaps indicating preparation for potential drug use. Alkaloids can sometimes be altered using heat so parching can be reasonably suspected to indicate preparation for some type of human use. It does not prove
It is not clear how many of these finds were in funerary context as was the case for the string of peyote and how many were found in residences (suggesting their employment by living people). The context can be complex as it is clear that burials occurred that intruded into older residential remains.
Their hypothesis is intriguing though; further work should be done to evaluate it even if it turns out to be a blind path.
A systematic review of the contents of rock shelter and cave excavations in the Chihuahuan Desert might prove a valuable avenue for anthropological studies of the religious and spiritual beliefs of people in this area before the invasion and occupation. The creation of small painted rocks and large pictoglyphs featuring shamanic themes are among the unique elements left by the archaic people who once lived around the mouth of the Pecos River.
A fascinating analysis was performed, by Bruhn & workers, on the aforementioned specimen of Cuatro Ciénegas peyote (the string of buttons) which was found by Taylor in 1941.
Bruhn et al. 1978 found 2.25% alkaloid in the Cuatro Ciénegas’ material in spite of it being thought to be from around 810 to 1070 AD. Their analysis was reported to show it to contain mescaline, lophophorine, anhalonine, pellotine and anhalonidine present in measurable amounts.
Phenolic alkaloids formed 35% of the total. It was noted that this is substantially lower than the 8% total alkaloid and 64% phenolic fraction which they observed in recently prepared peyote buttons.
They used tlc and GC (both with known reference samples) to determine this. See Bruhn et al. 1978 for more details.
A peculiar assessment was more recently presented by Bruhn et al. 2002, also DeSmet & Bruhn 2003 and also El-Seedi et al. 2005 where it was being noted that the total alkaloid content was lower in the far older Witte material from the Shumla Caves. They then went on to assert that while only 2% alkaloid was present there was no alkaloid other than mescaline that was detectable. That was intriguing enough but was not the end.
Bruhn then made the peculiar proposal that the reason they could only detect mescaline could be due to mescaline being more stable than the other peyote alkaloids. Reasonable enough on the surface yet it is noteworthy that in this Bruhn failed to mention any of his own previous work that contradicts this claim. Clearly something is in need of closer inspection. This claim about the Shumla specimens needs consideration in the light of his 1978 Cuatro Cienegas results. There was also his earlier report concerning degradation of peyote alkaloids in the study of 87 year old peyote buttons in Bruhn & Holmstedt 1974. In particular, their determination clearly showing that mescaline content apparently decreased *ahead* of the rate of decomposition of many of the other alkaloids. The cherry-picked selective presentation of facts in their 2002/2003/2005 account appear to have been overlooked by Bruhn’s peer reviewers or at least they dropped the ball in terms of what peer review is meant to accomplish. That is all actually fairly however when trivial compared to what they AND Bruhn’s crew missed.
The most interesting aspect of this so-called ‘mummified’ material in my mind is that, unlike Bruhn, Furst or Taylor, Terry & coworkers recognized that the specimens were not dried peyote buttons at all but rather were manufactured effigies of peyote created from some type of doughy material. (See images of both the Shumla and Cuatro Cienegas materials in Terry et al. 2006.)

Shumla peyote effigy
There are only the partial and hollowed out remains of three of these effigies are left due to the destructions of the rest of the materials during chemical analysis and radiocarbon dating. They were formed as a mixture of cactus materials combined with some sort of unidentified fibrous noncactaceous plant material and the resulting dough shaped by hand to vaguely resemble a living peyote cactus top. There was apparently at least two different makers of these artifacts over a period of some years as both their compositions and their dates varied. The most recently manufactured effigy had used entirely cactaceous materials.
More remarkable would be the assertion by Bruhn & coworkers that was just mentioned that this material contained 2% mescaline by weight *and* that only mescaline was detectable. Intriguing if true but in light of Bruhn & Holmsted previously establishing the deterioration of mescaline over time occurred at a faster rate than the deterioration of most of the other alkaloids I am left with far more questions than answers. It is obvious that we need to remain skeptical of Bruhn’s claim, no matter how intriguing it may be, as we proceed.
One thing that is clear, these ‘buttons’ were clearly manufactured artifacts. To be able to retain 2% mescaline after 5 millennia would require it to have once have had a far higher alkaloid content. The presence of ONLY mescaline would not be a result of some magical selective degradation process omitting mescaline but would absolutely require the involvement of some type of purification process. Or perhaps there are missing elements of this tantalizing puzzle that are presently defeating our understanding of exactly what has been established in these studies.
Did these prehistoric people really know of a route for purifying or at least greatly concentrating mescaline? It would certainly be interesting if true but establishing that Bruhn’s analytical results were actually valid has to be the starting point for answering this peculiar question and the burden of proof for establishing their validity is on the authors of that work.
Were these potent effigies a prepared drug form that were created for sacramental use rather than for simple use as effigies? No matter how they were prepared and no matter what their alkaloid content, they are clearly both deliberate and sophisticated in their preparation.
Or are Bruhn/El-Seedi/DeSmet’s reported analytical results simply needing to be questioned?
“Petrified” peyote buttons
The first reference to ‘petrified peyote’ was a misnomer in reference to the Shumla Caves’ adulterated & reconstituted 5 millennia old peyote effigies that were mentioned above. (See Terry et al. 2006 for details.)
More recently ‘petrified peyote buttons’ have been offered for sale (and finding at least one buyer) at a large southwestern Gem & Mineral Show and probably elsewhere.

Sold as petrified peyote button (front)

Sold as a petrified peyote button (back)
These do appear on first glance to vaguely resemble dried peyote buttons but are without a doubt either an agate or another form of chalcedony with a fine drusy quartz coating on one side. They lack the critical features (such as ribs, the distinctive apex and areoles) that are typically found in peyote buttons (see below).

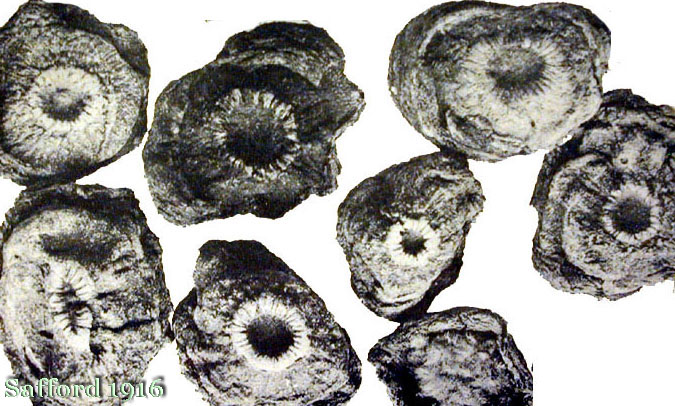
peyote buttons
They are amazing natural treasures but clearly are not of botanical origin.





















































































































































































































































{kind=link}